ESSEX BOTANY AND MYCOLOGY GROUPS
|
|
|
|
|
|
|
|
|
|
|
ESSEX BOTANY AND MYCOLOGY GROUPS |
|||||||||
|
Water poplars Essex txt:
|
The status and clonal distribution of Water Poplars, Populus nigra betulifolia in Essex and the features distinguishing them morphologically from the numerous exotic poplars now grown in the county. KEN ADAMS 63 Wroths Path, Baldwins Hill, Loughton. Essex IG101SH [Essex Naturalist 2009/2010] As part of a personal project to map the distribution of all the Water Poplar clones across southern England, all the isolated individuals and clusters of mature trees known in Essex have been sampled and their microsatellite DNA status determined for me by Forest Research, Roslin. Edinburgh. The distribution of the five male and two female clones detected in the county is presented here with a preliminary assessment of their wider distribution elsewhere in the region and some implications for their associated social history. Introduction What is a Water Poplar? It is the old English name (Cook 1676) for what has more recently been called the ‘Native’ Black Poplar or the ‘Atlantic race of the European Black Poplar’: Populus nigra betulifolia. Neither vernacular is particularly appropriate as it would seem that many if not all our trees are probably no more native than the so-called ‘English’ Elm; now known to have been introduced to England around the first century AD and to derive from an Italian clone (Gil et al 2004). Furthermore, Populus nigra extends right across Europe into central Asia and south into N. Africa, so it’s hardly appropriate to call P. nigra sensu lato the ‘European’ Black Poplar. Hence my endeavour to promote the old English name to minimise confusion with so-called ‘Italian’ Black Poplars and other Euroamerican and Euroasian/Hybrid/ Black Poplars and the various American Cottonwoods that are now grown in Britain. All poplar taxa are single sex trees, and they retain their sexuality if grown from cuttings. If you are just interested in the clone results, then jump to the section on Essex clones, as I am first of all going to describe and discuss the problem of distinguishing Water Poplars from some of the similar alien trees that are now planted in vast numbers in Essex. Our U.K. trees are all said to belong to a doubtfully distinct form separated off as var. or subsp. betulifolia. Although samples were collected for herbaria as long ago as the 1600s, and are now in the BM herbarium, nobody seems to have noticed any difference between our trees and those elsewhere in Europe until they were described by Francois Michaux, who found it growing on the banks of the Hudson river near Albany New York, and as large trees in New York City (1803), and thought it probably native to somewhere in the USA. It was subsequently propagated and sold by a firm in Rochester N.Y. under the name of P. elegans. It is now suspected that it was actually introduced to the U.S.A. from Europe some time in the 1700s. Even in Flora Europeae Edition One 1964, Flora of the British Isles 3rd revised edition 1989, and Oliver Rackham’s various books, there are no mentions of varieties other than the Lombardy Poplar. Widespread awareness of the existence of var. betulifolia as a distinct form seems to be very recent and to coincide with the growing desire to label taxa as ‘rare and endangered’ – much more justifiable if it’s not lumped in with a tree found right across Europe and into central Asia and N. Africa. In ‘typical’ betulifolia the laminas of the leaves and first soft shoots are pubescent when young, as are the petioles, which have more persistent hairs. The degree of hairiness of the petioles in U.K. clones is however very variable. Some are villous to the naked eye, others have tiny 0.5mm hairs only to be seen with a x20 lens, and some have no hairs at all – the hairs tend to snap off by late summer anyway. The axes of both the male and female catkins are also ‘typically’ hairy, - but here again forms have been described from the U.K. that have glabrous catkin axes combined with hairy petioles, as long ago as by Elwes & Henry 1913. As one moves south and east on the continent into drier climates the degree of hairiness of the leaves, petioles and first year twigs apparently increases, presumably as a xerophytic adaptation to stabilise the shearing layer, and another variety var. caudina Ten. (= var. pubescens Parl) Woolly Black Poplar - has thus been recognised. Whereas betulifolia occurs primarily in Britain, the low countries, and France from Normandy south to the Pyrenees; caudina occurs in Spain, North Africa, central and Southern Italy, the Balkans and Iran. The closely related var. afghanica (= thevestina) regarded by some as a separate species, and by others as a cultivar, has a smooth grey/white bark and pyramidal habit, and originated in central Asia but is now widely planted in southern Italy, N. Africa, western Asia and the Near East. It’s probably not hardy enough to grow over here. One of its cultivars: ‘Hamoui’ from Syria, is widely used for carpentry. [None of these hairy forms however, have the ciliate laminal margins of the American P. deltoides]. At the other extreme, the so-called var. typica or P. nigra var. nigra is completely glabrous. It is found in central and eastern Europe, and is said to be occasionally ‘planted’ in the U.K as opposed to occurring in the ‘wild’. The big question that arises however, is - are we simply talking about relative degrees of hairiness, or are there other characters which separate var. betulifolia, var. typica and var. caudina? If not then are we making a mountain out of a molehill? Elwes and Henry 1913 made an attempt to characterise betulifolia and typica. They say that the leaves of the two are identical, apart from the pubescence of the former, and the buds are greenish tinged with brown in the former and reddish in the latter, the twigs orange-yellow by the second year in the former and ash grey in the latter. They also mention, as many of us have found, that the degree of burring is very variable, as is the occurrence of epicormic shoots, - and I wonder if this could perhaps be caused by a widespread virus or phytoplasmid infection that man has selectively propagated in the majority of our U.K. material. As one progresses south in Europe the degree of variation of P. nigra increases, and is reflected in the number of clones, which are around ten times the number south of the Pyrenees, in Italy and the Balkans, as in the north of Europe (Cottrell et al 2005), the latter apparently having been recolonized after the last ice age by a small selection of clones from these presumed glacial refugia after they had struggled back across the barriers of the Pyrenees and the Alps. Fastigiate (tall and spindly) P. nigra clones. Before discussing how to distinguish Water Poplars in the field from the numerous hybrids involving hybridization at the specific level in Essex, we need to consider how to separate them from some of the widespread cultivars and hybrids within the P. nigra complex. The commonest is P. nigra ‘Italica’ (formerly called var. italica the Lombardy Poplar). [The modern convention is to enclose cultivar names in inverted commas and give them a capital first letter and regular as opposed to italic type]. This is probably a single gene mutant of P. nigra typica which has a dominant apical shoot and steeply ascending branches forming a narrow spire-like tree, the so-called ‘fastigiate habit’. It is a male clone that has been propagated by cuttings all over the world as an ornamental tree and wind break. It often has a short life, growing to full height in about 15 years. Because it is otherwise a typical P. nigra it has all the other characteristics of P. nigra, including insect parasites and diseases. Because typica is glabrous it too is glabrous. However there are complications. It has been crossed with P. nigra betulifolia to produce a whole family of male only cultivars lumped together as ‘Plantierensis’ – these are less spindly, have a heavy tapering trunk at the base, and are better adapted to the cool humid climate of northern Europe - but because betulifolia is involved they are hairy! Why mention it? Well they are now more commonly planted in the U.K. than ‘Italica’. Furthermore there are female fastigiated clones of P. nigra. One of these, quite commonly grown in the UK, is broader than ‘Italica’ and and resembles ‘Plantierensis’ – it’s called ‘Gigantea’. Both of these occur in Essex and have led to confusion with Water Poplars. There are several additional female fastigiate clones that have been selected and widely planted in France and around the Mediterranean and will probably appear over here. Why mention all these fastigiates? Several reasons. Firstly, it can be difficult to separate very young typica or betulifolia from one of the young fastigiates and secondly if one of the fastigiates has been coppiced it may have developed a broad habit. Thirdly, you will now appreciate that finding a young tree with hairy petioles is no guarantee that it is betulifolia. Fourthly, although these fastigiated poplars are all 99.9%+ P. nigra in origin they are so abundant that they are likely to genetically contaminate any seed produced by our betulifolia with a fastigiate habit. Finally – beware – a superficially similar ornamental fastigiate mutant of Common Oak Quercus robur, is all the rage with local authorities at the present time. A close look will reveal typical Q. robur leaves. It has for example been planted as a screen around the car park at the Hanningfield Reservoir Fishing Lodge. EuroAmerican/Asian Hybrids. In the OECD 2006 report it is stated that there are at least 30 recognized taxa of the genus Populus worldwide worthy of specific rank. These estimates apparently largely ignore the monumental tome, the ongoing web based multinational-authored e-Flora of China, in which are described and illustrated 73 species, and nine hybrids – as well as many varieties. Of these 43 species are additional to the 30 on the previously recognized list giving us around 73 species. This is in contrast to just 14 species in the whole of N. America. It would seem therefore that China is probably the centre of speciation of the modern genus, although Populus as a genus is believed to have its origins in the Palaeocene of tropical N. America. The southernmost naturally distributed species appears to be the Kenyan endemic P. ilicifolia. If you are confused already, the bewildering range of hybrids involving up to five generations of crossing/back crossing will further confuse. Many of them are naturally interfertile, and of those that are not mankind is doing his best to cross artificially! Not only have many of the species been further selected for ‘promising’ cultivar clones, numerous further clones have been and are still being selected from complex hybrid crosses. Genetic engineering is also now being applied to poplars, the hybrid between our P. tremula Aspen, and the American P. tremuloides for example has been genetically modified to remove the yellow colour that otherwise discolours pulp for paper making. The genus has been divided up into 5-6 subgenera of which the Tacamahaca (Balsam Poplars) and the Aigeiros (Cottonwoods and Black Poplars) cause us most concern in the U.K. These are all cross-compatible, and very closely related. Confusingly, if you put betulifolia shoots in a polythene bag for a few hours, or stand down-breeze of a freshly leafing tree in spring, you will detect a strong balsam smell that emanates from the sticky buds. In the wild all of these two groups are essentially riparian species characteristic of river flood plains, but they are poor competitors, and require open weed free conditions for seedling establishment. The hybrid P. nigra x P. deltoides (P. x canadensis) or (P. x euramericana) confusingly called Italian Black Poplar (because the first cross arose in Italy) or Hybrid Black Poplar (even more ambiguous) has now been cloned in numerous forms, all faster growing than P. nigra and now planted in their thousands in Essex. Curiously the potential cross P. nigra female x P. deltoides male is incompatible, and only works the other way round. In Table One I have expanded the ancestry of some of the commoner male and female hybrid cultivars grown in the U.K. covered in Stace 2010 to give some idea of the complexity: TABLE ONE:
All these have the common characteristic that they are glabrous, giving us one way of separating them from betulifolia. All that is except ‘Marilandica’ which has downy petioles (where did it inherit this character from?) Just to complicate matters however, a cross between betulifolia and deltoides – [and therefore pubescent] has recently been re-introduced into the UK from America. Furthermore, although ‘Serotina’, ‘Marilandica’ and ‘Regenerata’ are largely used in amenity/ornamental plantings, a further hybrid ‘Robusta’ with a tall straight trunk is planted in rectangular arrays in damp ‘Cricket-bat Willow-like’ plantations for timber, and in long rows as screens or wind breaks, and is probably the most abundant form in south-east England. Because it has betulifolia as one of its ancestors it is pubescent. ‘Eugenii’ is also grown in plantations, now more commonly than ‘Robusta’ and also has the habit of a Cricket-bat Willow, but it is glabrous.
All the Poplars have a base number of chromosomes of n=19, and most of them are diploid with 2n=38, and this applies to the hybrids involving nigra and deltoides, which appear to be fully fertile and able to backcross with each other. P. nigra specific aphids. So how can we be certain we have a tree of Populus nigra betulifolia/typica in our sights? Even with experience it is often difficult to be sure without DNA fingerprinting. One way of getting a favourable indication is to look for the presence of P. nigra specific aphid parasites, in particular the spiral gall forming aphid Pemphigus spyrothecae. P. spyrothecae does not have a secondary host. It emerges in spring and in mid-late April/early May begins to feed on a young emerging leaf, puncturing the petiole in a helical line which induces the tissue to swell, forming a groove around the aphid and to twist into a spiral gall of 2-3 complete turns over a period of 6-8 weeks. Inside, the aphid gives birth to 20-30 wingless aphids in June, and then a second generation of winged aphids emerges from the gall in August, and these live in the bark where they lay eggs. These hatch into males and females which copulate and lay over-wintering eggs on the bark. The following aphids are regarded as specific to the P. nigra aggregate: TABLE TWO:
P. spyrothecae is usually the most abundant of the P. nigra specific gall causers, and occurs on betulifolia/typica, ‘Italica’ and the ‘Plantierensis’ complex. It does not occur on ‘Serotina’, ‘Marilandica’ or ‘Robusta’, - but it does occur at low frequency on some of the trees that I am doubtful about, and suspect might be deltoides-based hybrids back-crossed with P. nigra. When I get a ‘phone call or e-mail from the other side of the county asking me to check out a WP I first of all ask if it has any spiral galls on the petioles. This is not an infallible indication of betulifolia because they are equally abundant on typica, ‘Italica’ and ‘Plantierensis’ but it does normally rule out a contribution from deltoides. If it is one of the P. nigra group, the spiral galls are usually very abundant, commonly as many as 2 and up to 6 per petiole. Leaves with spiral galls usually drop first in the autumn, and can usually be found recognizably in the leaf litter throughout the winter. I should point out that P. spyrothecae will not attack trees that are water stressed. It is possible to find a tree smothered one year and the next either none at all, or only literally a handful, if the tree is suffering from drought. A further indication of drought symptoms is leaf size. A heavily pollarded or crown reduced tree may produce very small leaves for several years during the recovery phase, and may take p to five years before it recovers the ability to produce flowers. Looking at the other possible P. nigra faithful aphids, I have never seen P. protospirae, not surprising perhaps if it has Berula and Sium as its secondary hosts. Maybe it will turn up in the Cambridge or Norfolk fens. It has more than 3 turns to its spiral gall. DIAGRAM ONE |

|
|
Common gall inducing aphids specific to Populus nigra sensu lato. Right to left: Pemphigus spyrothecae, P. bursarius, P. populi & P. populinigrae agg. P. bursarius, the Pouch Gall is another petiole specialist and is the next in frequency. It occurs commonly on ‘Italica’ and occasionally on betulifolia. P. populi, which forms a globular gall on the upper surface of the midrib with an access hole below and often splits open above, is less common. The next three all form similar linear pouch-like galls on the mid-rib, and you have to look at the aphids inside to identity which is which. Something I have not yet done although the pouches which start off green and then go red are not uncommon Characteristics of P. nigra versus P. x canadensis [ = euroamericana] Maiden P. nigra trees that have been allowed to grow for 60 years or more are now extremely rare, most trees are either low, middle or high pollards. In towns and parks ornamental P. nigra often gets treated like a Hybrid Lime or London Plane. If they are regularly mop-head pollarded most withstand it O.K. but if significant crown reduction or pollarding is carried out on a tree that has missed several cycles the exercise is frequently fatal. Many modern plantings are ridiculously over-crowded, in avenues or tightly packed groups. For these reasons it is virtually impossible to discern from the form of the tree whether you have a betulifolia/typica. Quite often one of the fastigiates has at some time been coppiced and it then becomes very difficult to separate them from betulifolia. I have listed some characteristics separating the commoner deltoides/nigra hybrids from nigra typica/betulfolia: 1. Populus x canadensis (Populus nigra typica x P. deltoides) ‘Italian’ Black Poplar Leaves with cartilaginous margins, thin, lamina not leathery, light green, rather large, teeth rounded, hooked and curving forwards towards the apex, base truncate to cordate and in some forms cuneate, two glands at junction of petiole and lamina, petiole and leaf veins glabrous. Terminal shoot leaves similar to the others. Buds and young shoots not smelling of balsam. Trunk and branches without burring or bosses. Branches ascending, not significantly downward arching. Bark deeply fissured with rounded spaghetti-like segments, usually grey in colour. Commonly supports Mistletoe. Never supports the aphids Pemphigus spyrothecae, P. bursarius, P. populi or P. populinigrae agg. 2. Populus nigra betulifolia/typica Water Poplar, ‘European’ Black Poplar, ‘Native’ Black Poplar. Leaves without cartilagenous margins, lamina rather thick and leathery, deep green, margin sinuous around most of circumference, may have slightly hooked teeth towards the base. Base of mature leaves cuneate to truncate but never cordate. No glands at junction of petiole and lamina initially but may be one later on, petiole and main veins below softly hairy in spring in betulifolia. Terminal shoot leaves markedly diamond shaped with cuneate base and long pointed apex. Further back, leaves may be truncate but not cordate. Buds and young shoots smelling of balsam. Trunk and branches usually, but not always, burred and bossed with epicormic shoots. Trunk black, fissured with slit-like fissures as if cut by a knife and pulled apart. Upper branches may however have grey spaghetti-like bark in large mature trees. Main branches arching over in wide arcs, often touching the ground. N.B. in recently pollarded trees these may be steeply ascending for the first 5-6 years. Rarely if ever supports Mistletoe [2 records]. The aphids Pemphigus spyrothecae, P. bursarius or P. populinigrae agg. all specific to P. nigra (including P. nigra typica, P. nigra ‘Italica’ (Lombardy), P. nigra betulifolia and P. x plantierensis). DIAGRAM TWO |
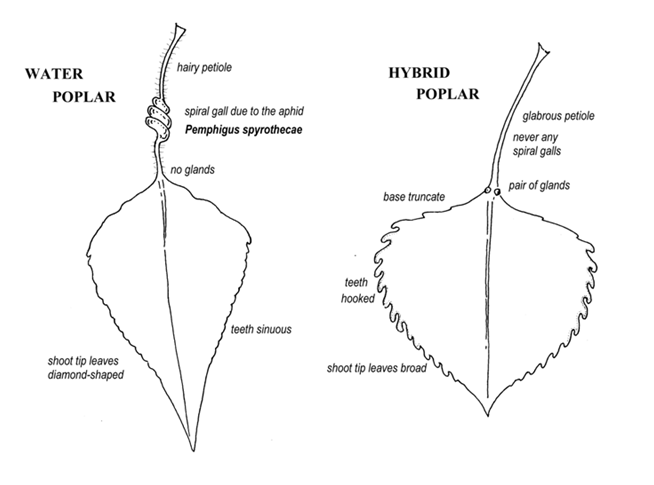 |
|
Schematic of the characters distinguishing a Water Poplar from the various hybrid clones involving the American Eastern Cottonwood P. deltoides and P. nigra typica. Given these possibilities, how do we go about separating P. nigra in the field from first generation hybrid black poplars and possible second generation back-crosses? First of all I should mention that there are a lot of red herrings in the literature. Supplementary characters such as bronzing of young leaves or red pigmented petioles, for example seem to be reactions to a high U.V. flux. Since P. nigra usually leafs up before the hybrids in the spring, it may not exhibit either characteristic, whereas the latter leafing up later on when the UV flux is higher usually do. However, if we have fine sunny weather in April/May P. nigra can be just as bronzed or red-petioled, though they may not be the following year. They all have red male catkins, but those of ‘Robusta’ emerge first and are dark red, betulifolia male catkins emerge next, and are of such a bright red colour that the whole tree can be seen glowing red for a mile or so away. Next come the other various hybrids, mostly of a darker less conspicuous red, not so readily picked out from a distance. [N.B. male catkins drop soon after anthesis but their shrivelled remains can also usually be found in the leaf litter or grass under a tree well into August. Similarly, the remains of the female husks and fragments of fluff can usually be found well into the autumn, thus enabling the determination of the sex of a tree well after the flowers have disappeared on the trees.] In addition to all the above trees, we also have other Poplar taxa in Essex. The Aspen, Populus tremula, the White Poplar Populus alba, and their hybrid Grey Popular, Populus x canescens; and the true Balsam Poplars from N. America: Populus trichocarpa and P. balsamifera, the Western and Eastern Balsam Poplars – and not forgetting their hybrids! Why the interest in Water Poplar clones? There has been a growing realization Europe wide, that enormous quantities of pollen are being produced by the tens of thousands of ornamental and commercial plantings of crosses between the exotic American Eastern Cottonwood P. deltoides and P. nigra typica; and P. nigra betulifolia; together with fastigiate clones of Populus nigra typica, in particular the male ‘Italica’, and the various male P. nigra betulifolia x ‘Italica’ clones known collectively as ‘Plantierensis’. Although females of P. nigra have been shown to respond preferentially to conspecific pollen in mixed pollen clouds (Vanden Broecka et al 2006), if none alights on their stigmas, they will accept pollen from, and cross with P. x canadensis. They can also cross with ‘Italica’, and Water Poplar males can cross with the fastigiate female ‘Gigantea’. This is not a significant problem in the U.K. however, as suitable conditions for germination and establishment no longer exist due to destruction of our riparian forests. It is however, a problem on the continent, where for example around 50% of the young trees along the Rhine were found to be of hybrid origin (Smulders et al 2008), and will become our problem if riparian forests are re-established in the U.K as some counties are planning. In recent decades, enthusiasts have also become concerned about the apparent scarcity of Water Poplars, and are now aware of the need to conserve as much remaining genetic variation as possible, to try and avoid the sort of epidemic disease outbreak that decimated our monoclonal Ulmus procera. This has led a number of counties to extend and refine the recording of the exact location and sex of individual trees started by the late Edgar Milne-Redhead, and they have begun to collect cuttings from every individual tree, and one or two from each group of trees, and to propagate them in clone banks. Recognition of individual clones using DNA fingerprinting with microsatellites has made it possible to avoid duplicating identical clones unnecessarily, and to make sure that as much genetic variation as possible is collected. Assignment of Clone numbers using DNA microsatellites. Previous attempts to use the variation in DNA sequences for characterizing individual trees, by using enzymes to chop up the DNA where particular runs of nucleotide bases recognized by the enzymes occur, were analysed by determining the relative lengths of the fragments that are produced. Thus if the sequence of one of these sites changes it will not be cut by the enzyme and the fragments either side of it will be joined together to produce a larger fragment. Alternatively if a mutation generates a new cut site somewhere, a fragment will be cut into two smaller fragments. These techniques are however not very discriminating, and are unable to pick up differences between closely related individual trees with any reliability. The technique now in use at Forest Research, Roslin however, uses sites in the DNA called microsatellites. These are small segments of highly repetitive DNA that are more liable to repetition mutations during replication than other DNA segments. By using seven of these in different parts of the genome, it becomes possible to pick up differences between individual trees, the microsatellites in the offspring grown from seed being a combination of those of the parents. These sequences are however stable over at least several generations so that if trees are grown from cuttings the patterns are identical with the tree the cuttings are taken from. Thus it becomes possible to ‘fingerprint’ individual vegetative clones over several generations. What is becoming apparent from the studies thus far is that virtually all our P. nigra betulifolia trees have been planted by man as cuttings derived from a relatively small number of original trees. Trees that are derived from another by cuttings are said to be vegetative clones of that tree, and we have found that in Britain some of the clones are now represented by thousands of trees. In addition to the seven microsatellites common to the Aigeiros (Cottonwoods and Black Poplars) subgenus, there is a shared gene that in P. deltoides has a sizeable chunk of DNA missing – a DNA deletion, that is being used by Forest Research to detect first generation P. x canadensis hybrids. Unfortunately, in a backcross between such a hybrid and P. nigra, since the deletion is only on one chromosome of a homologous pair, there is a 50% chance that it will be lost at meiosis. It is not therefore possible at the present time to detect such backcrosses infallibly, although in the future the Forest Research team are planning to use additional chromosomal markers. For the same reason, any presumptive deltoides gene that might account for its avoidance by our European P. nigra specific aphids may also be lost in a backcross thus possibly accounting for what appear to be deltoides backcrosses having some spiral galls. As the fingerprinting of trees is an expensive operation, although all lone trees have been fingerprinted, in the case of a row or a cluster of trees at one site, if they look identical, are the same sex and appear to be the same age, only two of them have been sampled, and if these are the same it has been assumed that they are all the same clone. The trees have been sampled in two phases. Initially all the Essex tree samples collected by Peter Ennis and myself and used to establish stool beds in the clone bank at Daws Halls were analysed by Forest Research. Then in 2009 I collected samples from the all the remaining unsampled lone trees and clusters, making it possible to assign clone numbers to a total of 236 trees. The actual number of trees alive today is in some cases less than those quoted below, as several of them have fallen or been cut down since the survey began in 1997. CLONE 28. This male clone is the most abundant in Watsonian Essex ,Vc18 & 19, (that is including the London Boroughs west to the R. Lee). Assuming that all trees of the same apparent vintage growing in clusters are the same clone as samples taken from a few of them, they total at least 122 trees. This clone is identical to, and has therefore been propagated vegetatively from, the same parent as all the 7,000+ trees originally planted around the Manchester conurbations from about 1850 onwards, and in numerous sites across southern England including London. In Vc 18 this includes the hotly contested tree in the Lee Valley Park beside the Waltham Park estate, and the three trees at Town Mead, Scouts site; Watham Abbey; all the trees in Hainault Forest Country Park (at least 35 originally); the lone tree in Epping Forest, just west of Ching Brook; the six trees originally planted in Hospital Wood, Claybury; the two planted in a hedge south of Chigwell School playing fields; the eight planted around the old sewage works ruins by the Roding south of Buckhurst Hill School; the lone tree in the car park at St. Margaret’s Hospital, Epping; the seven trees in the grounds of the Victorian Abbey Mill sewage pumping station at Stratford, together with the very stunted lone tree just over the fence by Abbey Creek; the three trees in a hedge across Walthamstow Marshes; the two trees on the east bank of the Lee navigation on Leyton Marshes [together with some 67 trees of the same clone on the Middx. side of the Lee on Hackney Marsh Recreation Ground]; the one tree in the recreation ground at the northern end of Lechmere Avenue, Woodford Bridge; the three trees on the west boundary of Upminster Junior School; the huge maiden tree at the western end of Corbets Tey, Upminster; two small trees recently planted in the SE corner of the Great Warley Hall interchange on the A127; the at least three trees originally planted by the industrial estate south of the A13 at Aveley; the large forked tree by Brentwood Road, Bulphan; the two (now one) by the Rose Pub. Shenfield; the one in the garden of 122 Chelmsford Road, Shenfield; the tree on the corner of Roberts Road and Devonshire Road, and the one in Devonshire Road; Laindon. A young newly planted tree inside Coalhouse Fort, East Tilbury, and the three trees by the entrance to Linford Schools; one tree in the Cliff Gardens below Wilson Road, Clifftown, and three trees in Southchurch Hall Gardens; Southend. The two trees on the Beckton Sewage works site, lost when the shaft was constructed down to the new Tideway Tunnel, were probably also clone 28. And the tree behind a house in Victoria Road, Laindon is almost certainly this clone. In Vc 19, clone 28 includes the lone tree at the back of the vicarage at Epping Green; the seven trees at Glen Faber; two trees planted at Woodhill, Harlow New Town; two trees by the River Can, West Park, and the single tree by the viaduct in Central Park; Chelmsford. The four trees among a row of hybrids along the north boundary fence of Carver Barracks, Wimbish; the lone tree at Hall Farm, Gestingthorpe; the three trees in the front garden of Hillcrest, Horkesley Heath; the two trees originally at Rookery House, Great Horkesley; the three at Bounds Farm, Goldhanger; the lone tree in Brook Road, Tolleshunt Knights; and the tree in the middle of Osea Island, just west of Meeting Lane. CLONE 47. This female clone is confined in Essex to Vc19, and is only known so far otherwise to occur at one site in west Suffolk Vc.26 and on the Hertfordshire border, although we expect to find it elsewhere in Hertfordshire and in Cambridgeshire in future sampling. The biggest tree found so far is a huge maiden by the Pincey Brook just south of Lang Bridge at Hatfield Broad Oak. It may possibly be the original source of the rest of the population in the west Essex area. There are, however some very large and very old pollards in the Roydon area centred along a stream at World’s End. There were originally eight large pollards along the stream, but these have now been extensively cloned and planted at various sites in the Roydon area and on East Farm in Harlow. A further ten old pollards occur east of Tyler’s Cross, one along Pardon Brook, and nine along a ditch flowing into it; and eight more occur along the R. Stort north of Roydon station in what is now part of Hertfordshire. In addition there are two large pollards of this clone just west of Abbess Roding church; and two more along Little Hallingbury Brook at Gaston Common, and there is an isolated tree to the west of Wall Wood at Woodside Green, now in a very sorry state. In all there are around 30 veteran trees in Vc19, but large numbers of new trees of this clone have been planted in the area around Roydon. CLONE 25. This male clone is the next in terms of numbers of veteran trees [24] in the county. It appears mainly to be a Suffolk clone, with a concentration of occurrences in southern Suffolk spilling over into Vc19. It has however turned up in Leicestershire, and at least one has been planted recently in Richmond Park. There is a single tree at Ridgewell, between the Colne and the Stour; one at Northey’s Farm, and another at Hill Farm; Gestingthorpe. There are two at Hickford Hill, Belchamp St. Paul; one at Lapwing Meadow, Belchamp Walter; one at Whitehouse Farm, Belchamp Otten; one at Hole Farm Bulmer; and one at Liston Weir; there are three at Aylett’s Farm Halstead; there are eight in a hedge at Rookwoods, Sible Hedingham; and then there are four trees in west Essex, with one at Upper Pond Street, Elmdon; two at Tye Green in Harlow; and finally one at Rose Farm, Epping Upland. There are no veterans of this clone that I am aware of in Vc18. CLONE 32. A female clone this one, with 22 veterans represented in Essex, again largely a Suffolk tree, this time mainly in East Suffolk Vc.25, but it also occurs in Leicestershire, Cheshire, Middlesborough and as three veterans in Richmond Park. Apart from 4 large trees in Dovercourt Cementery, rather than spilling over the border into Vc.19 however, it seems to have been planted at isolated sites right across Essex. A single tree by the R. Blackwater just north of Blue Mills, Wickham Bishops (which has now been extensively propagated in the Witham area); three trees, originally in a cluster by the R. Chelmer just down stream of Little Waltham village; three trees in a roadside hedge in Chickney just east of Sibleys; and most strange of all, a tree in a garden at Carpenters, High Ongar; nine trees originally in a line in Eastbrook End Country Park by the Beam River, in Dagenham (now propagated elsewhere along the valley and allegedly in Buckingham Palace garden); two more veterans inside Coalhouse Fort, East Tilbury; one in Jones Memorial Park, Sutton at Southend (now gone); one in Chalkwell Park; and strangely, one in the stackyard at Doggetts Farm, Rochford. The same clone has also turned up as an isolated tree in Russell Square in the centre of London; a single tree at Putney Heath; and in Footscray Churchyard in west Kent Vc.16, from where it has been extensively propagated in the adjoining meadows. CLONE 23. With 15 veterans in Essex this male is the next most abundant clone. It is well represented in Suffolk and spills over the border into Vc.19 with three trees originally on an island at Stratford St. Mary on the Stour in Langham; one at Broom Knolls, Lawford; one at Boxted Mill and three others by the R. Stour at Boxted; one at Foxearth Mill on another island in the R. Stour; one by Toppesfield Brook, beside the White Hart pub. at Great Yeldham; and then we have four trees in the Cliff Gardens at Chalkwell, and one isolated tree in Gunner’s Park at Shoebury; Southend. Elsewhere I have found a tree at Kew on the south bank of the Thames (believed to have been planted by Edgar Milne-Redhead); and three in the Cotswold Water Park in Wiltshire, which suggests it will turn out to be a widespread clone nationally. CLONE 34. Finally we have the strangest clone in terms of distribution. This clone is male, and in Essex is represented by about fifteen trees. It has been found as far away as Cheshire and Middlesborough, and yet forms a cluster in the Frinton/Holland area. Although we know that Edgar Milne-Redhead brought various Cheshire tree seedlings to Suffolk, these trees obviously pre-date that event, as there is a large veteran of this clone in a hedgerow just south of Hillside Close in Frinton, which seems most likely to be the source of cuttings for the at least seven trees originally planted around Frinton, and the one in a roadside hedge at Great Holland. There is also an isolated tree on the south bank of the R. Stour at Dedham; and there are four trees in Tolleshunt D’Arcy cemetery. Even more remarkable is the single isolated maiden beside the R. Roding at Curtismill Green that we used for the cover picture of our EFC Newsletter No: 58 – and which has now snapped off. A single tree has also turned up in Clapton Square, Hackney, and ten trees occur on the west bank of the R. Lee beside Spring Hill Park, Tottenham. NEW CLONES Two Essex clones have turned out to be new, both male, and have yet to be assigned a clone number. One of these occurs as just a single tree in a hedgerow west of Waples Mill, Abbess Roding; the other occurs in a hedgerow behind the garage at Jack’s Hatch, Roydon. Strangely, this tree has a duplicate on the west bank of the R. Lee in South Island Marina, Ponders End, Middx. OTHER TREE CLONES As I have already pointed out it is not always possible to be sure of the identity of a tree, particularly if it has been pollarded or coppiced, or if it is a backcross. Several such trees have had their identity at least partially resolved by the microsatellite results. I was suspicious about a female tree at Bishop’s Court, Margaretting, that had spiral galls, but did not have a burred trunk or hairy petioles. So much so that when asked I did push for a TPO, although I did not realize at the time that this would lead to its being cut down! The base produced suckers the following year however, and I was able to get it fingerprinted. It turned out to have some microsatellites characteristic of ‘Italica’, and is probably therefore one of the ‘Plantierensis’ complex. A large tree by Salter’s Meadow in the village of Tolleshunt D’Arcy has also turned out to have ‘Italica’ DNA in its makeup. A female tree in front of St. Clement’s Court in Leigh on Sea however, has a profile that includes some of the microsatellites found in the cultivar ‘Gigantea’. Similarly, two female trees (originally four) in a roadside hedge at Chignall St. James were reported to Edgar Milne-Redhead as betulifolia by the late Colin Ranson. Although they looked somewhat like betulifolia, I was suspicious as the leaves had glands at the apex of the petiole and the teeth were rather hooked. When tested they revealed that the DNA deletion unique to P. deltoides was present. Discussion One of the most intriguing aspects of these clonal distributions is the questions that arise about the human history behind the patterns. To some extent the relative size of the veterans, enables some indication of the chronology of propagation from a tree at one site to another in a local cluster, but the motivations for taking cuttings of particular trees and planting them in remote locations in other parts of the country are likely to be largely lost in the unrecorded history of individuals families. Maybe certain individuals reading about trees mentioned here that they are familiar with may be able to throw some light on some of these puzzles. Unfortunately, most of the trees will have been planted several human generations back, and the information is therefore probably lost in the mists of time. The widespread occurrence of Clone 28 is almost certainly due to a major marketing effort countrywide, in the latter half of the 19th and early part of the 20th C, probably as a supposedly pollution resistant clone. Others have probably been moved around, as individual farming families have moved around the country, to use them as a major source of home grown construction timber. References ADAMS, K. J. (2010). Progress with a project to locate and DNA fingerprint the Water Poplars of southern England. BSBI News. 114: 15-19. A’HARA, S., SAMUEL S. & COTTRELL, J. (2009). The role of DNA-fingerprinting in the conservation of Black Poplar. British Wildlife 21(2): 110-115. COOK, M. (1676). Manner of raising, ordering and improving forest trees, p. 81. London. COTTRELL, J. E., KRYSTUFEK, V., TABBENER, H.E., MILNER, A.D.., CONNOLLY, T., SING,L., FLUCH, S., BURG, K., LEFE’VRE, F., ACHARD,P., BORDA’CS, S., GEBHARDT, K., VORNAM, B., SMULDERS, M.J.M., VANDEN BROECK, A.H., VAN SLYCKEN, J., STORME, B., BOERJAN, W., CASTIGLIONE, S., ROSSATI, T., ALBA, N., AGU’NDEZ, D., MAESTRO, C., NOTIVOL, E.,BOVENSCHEN, J., VAN DAM, B.C. (2005). Postglacial migration of Populus nigra L.: lessons learnt from chloroplast DNA. Forest Ecology and Management. 219; 293–312. E-FLORA OF CHINA: www. eFloras.org
ELWES, J.H. & HENRY, A (1900-1913). Trees of Great Britain & Ireland. Privately published. GIL, L., FUENTES-UTRILLA, P., SOTO, A., CERVERA, M.T. & COLLARDA, C. (2004). English Elm is a 2,000-year-old Roman Clone. Nature 431: 1053. ORGANISATION FOR ECONOMIC CO-OPERATION & DEVELOPMENT (2006). Safety assessment of transgenic organisms. Vol:2. Section 4. 110-117. Consensus Document on the Biology of Populus L. (Poplars). Paris. France.
SMULDERS, M.J.M., COBBEN, M.M.P., ARENS, P., & VERBOOM, J. (2008). Natural hybridization between Populus nigra L. and P. x canadensis Moench. Hybrid offspring competes for niches along the Rhine river in the Netherlands. Tree Genetics and Genomes 4: 663-675. STACE, C. (2010). New Flora of the British Isles. 3rd edition. p.p.1231. Cambridge University Press. VANDEN BROECK, A.N., COTTRELL, J.E., QUATAERT, P.,BREYNE, P., STORME, V., BOERJAN, W. & VAN SLYCKEN, J. (2006). Paternity analysis of Populus nigra L. offspring in a Belgian plantation of native and exotic poplars. Ann.For.Sci. 63:783-790. Acknowledgements
The enthusiasm of Peter Ennis for Water Poplar recording and cloning got me bitten with the bug initially, and I must also thank all those who have reported possible Water Poplar trees in Essex over the years, in particular Bob Creber, Keith Turner, Mark Hanson, and Tim Pyner. And over the last three years I have much appreciated the patient cooperation of Stuart A’Hara, Joan Cottrell and Sam Samuel at Forest Research, Roslin, Edinburgh in analysing my samples and sorting out their clonal status using the Smulders et al microsatellite markers. Fingerprinting costs for the second phase of sampling were covered by the Essex Biodiversity Project.
|